

These notes are in three parts:
All indented paragraphs are quotes from In Search of Memory: The Emergence of a New Science of Mind by Eric R. Kandel published by Norton in 2007. (NB. I have emboldened some key concepts.)
The non-indeneted paragraphs contain my reflections on the quotes
Three exercises Penny Tompkins and I designed to explore the perception of space.
In his very readable book, Eric Kandel, Nobel prize winner for Medicine or Physiology, intertwines a combination of cognitive psychology, neuroscience and molecular biology with his own personal quest to understand memory. The following quotes have been selected because they relate to the neurobiology of space perception. They have been collected together into three sections which describe the role of:
- The pyramidal cells of the hippocampus
- Selective attention
- A synaptic strengthening mechanism called long-term potentiation.
1. The role of pyramidal cells of the hippocampus
In 1971, John O’Keefe at University College London made an amazing discovery about how the hippocampus processes sensory information. He found that neurons in the hippocampus of the rat register information not about a single sensory modality — sight, sound, touch, or pain — but about the space surrounding the animal, a modality that depends on information from several senses. He went on to show that the hippocampus of rats contains a representation — a map — of external space and that the units of that map are the pyramidal cells of the hippocampus, which processes information about place. In fact the pattern of action potentials in these neurons is so distinctly related to a particular area of space that O’Keefe referred to them as “place cells.” Soon after O’Keefe’s discovery, experiments with rodents showed that damage to the hippocampus severely compromises the animals’ ability to learn a task that relies on spatial information. This finding indicated that the spatial map plays a central role in spatial cognition, our awareness of the environment around us. (pp. 281-282)
As soon as the metaphor of “a map” is used it brings with it entailments of a ‘map reader’. But, according to current neuroscience, there is no map reader in the brain, just synchronised electrical and chemical reactions from which behaviour emerges. Thus the ‘map’ Kandel refers to is not a map in the ordinary sense of the word, it’s a configuration of neurons whose pattern of firing contributes to the animal being able to navigate round its environment.
[That] the spatial memory of environments has a prominent internal representation in the hippocampus is evident even anatomically. Birds in which spatial memory is particularly important — those that store food at a large number of sites, for example — have a larger hippocampus than other birds. London taxi drivers are another case in point. Functional magnetic resonance imaging [fMRI] revealed that after two years of [learning ‘The Knowledge’ of the] streets of London, taxi drivers have a larger hippocampus than other persons of the same age. Indeed, the size of their hippocampus continues to increase with time on the job. Moreover, brain-imaging studies show that the hippocampus is activated during imagined travel, when a taxi driver is asked to recall how to get to a particular destination. (pp. 305-306)
We believe that because much the same neurology is involved in imagining doing something and actually doing it, this explains one of the benefits of “making words physical”. David Grove used to emphasise the need to facilitate the client to shift from conceptual descriptions which have no coordinates to located symbolic representations. Then parts of the brain that know about space, movement and interactions in the physical world can be brought into service during the consideration of an abstract problem or desired outcome.
In all living creatures, from snails to people, knowledge of space is central to behavior. As John O’Keefe notes, “Space plays a role in all our behaviour. We live in it, move through it, explore it, defend it.” Space is not only a critical sense but a fascinating one because unlike other senses space is not analyzed by a specialized sensory organ. How, then, is space represented in the brain? Because we do not have a sensory organ dedicated to space, the representation of space is a quintessentially cognitive sensibility: it is the binding problem writ large. The brain must combine inputs from several different sensory modalities and then generate a complete internal representation that does not depend exclusively on any one input.
(pp. 307-308)
Interestingly, Kandel regards space as a “sense” that is “generated” by a synthesis of the more traditional senses.
The brain commonly represents information about space in many areas and many different ways, and the properties of each representation vary according to its purpose. For example, for some representations of space the brain typically uses egocentric coordinates (centered on the receiver), encoding, for example, where a light is relative to the fovea or where an odor or touch comes from with respect to the body. Egocentric representation is also used by people or monkeys for orientating to a sudden noise by making an eye movement to a particular location. For other behaviors, like memory for space in the mouse or in people, it is necessary to encode the organism’s position relative to the outside world and the relationship of external objects to one another. For these purposes the brain uses allocentric coordinates (centered on the world). (p. 308)
Modern Sat Navs also give us a choice to use two perspectives: From within the vehicle (route perspective) and from above the vehicle (survey perspective). In NLP (Neuro-Linguistic Programming), these perspectives are called ‘associated’ (first-position) and ‘dissociated’ (third-position).
These are similar but not exactly equivalent to the use of ego- and allo-centric coordinates. In particular, allocentric coordinates do not contain the ego whereas in a survey perspective the car, the “ego”, is shown on the screen.
From a Symbolic Modelling perspective it’s not quite so simple. Egocentric is actually own-body-centric. But because ‘a perceiver’ can move independent of the ego’s physical body, egocentric and allocentric coordinates can be used in a variety of combinations. In fact, we doubt the sole use of allocentric coordinates is possible. Even if a person is using a map “centred on the world” they still have to perceive the map from somewhere. That’s why it makes a big difference when you turn a map upside down — the map is still using allocentric coordinates but your egocentric perspective on those coordinates has changed.
Egocentric coordinates are common in Symbolic Modelling as a client’s metaphor landscape is usually centred on their body. However, once a landscape takes on an independence from its owner, it forms ‘the world’ upon which allocentric coordinates can be based. Then the landscape can remain fixed while attention moves around it using either type of coordinate. However, there is always an “ego”, a perceiver doing the perceiving from somewhere – even if that is from outside the client’s body.
Clean Space has the effect of turning a client’s egocentric coordinates into allocentric coordinates by “nailing their history to the floor so that it can be examined” as David Grove so graphically referred to it.
The spatial map discovered by O’Keefe differs radically from the egocentric sensory maps for touch and vision, because it is not dependent on any given sensory modality. O’Keefe found that as an animal walks around an enclosure, some place cells fire action potentials only when that animal moves into a particular location, while others fire when the animal moves to another place. The brain breaks down its surroundings into many small, overlapping areas, similar to a mosaic, each represented by activity in specific cells in the hippocampus. This internal map of space develops within minutes of the rat’s entrance into a new environment. (pp. 308-309)
An analogue in Clean Space is the breaking down of a complex abstract issue into several small areas, each represented by a verbal or nonverbal description of knowledge. What is new (to us, at least) is the notion of “overlapping” spaces — something we want to explore on the day.
Unlike vision, touch, or smell, which are prewired and based on a priori [pre-existing] knowledge, the spatial map presents us with a new type of representation, one based on a combination of a priori knowledge and learning. The general capability for forming spatial maps is built into mind, but the particular map is not. Unlike neurons in a sensory system, place cells are not switched on by sensory stimulation. Their collective activity represents the location where the animal thinks it is. (p. 309)
In some ways “forming a spatial maps” seems parallel language acquisition. A baby comes into the world ready to acquire language but which particular language they learn depends on where and with whom they live.
And “where the animal thinks it is” is not a single thought, but multiple perceptions which somehow emerge in consciousness as an apparently unified representation. Even
when you know a visual illusion is fooling your neurons, it doesn’t stop you ‘seeing’ it as real. The two types of knowledge are processed in different ways and in different parts of the nervous system.
2. The role of selective attention
Selective attention is widely recognized as a powerful factor in perception, action, and memory — in the unity of conscious experience. At any given moment, animals are inundated with a vast number of sensory stimuli, yet they pay attention to only one or a very small number of them, ignoring or suppressing the rest. The brain’s capacity for processing sensory information is more limited than its receptors’ capacity for measuring the environment. Attention therefore acts as a filter, selecting some objects for further processing. This focusing of the sensory apparatus is an essential feature of all perception. (p. 311)
Attention also allows us to bind the various components of a spatial image into a unified whole. [Kandel’s mice studies show that] even ambient attention (the attention that is present in the absence of stimulation) is sufficient to allow a spatial map to form and become stable for a few hours, but such a map becomes unstable after three to six hours. Long-term stability correlates strongly and systematically with the degree to which an animal is required to pay specific attention to its environment. Thus, when a mouse is forced to pay a lot of attention to a new environment, by having to learn a spatial task at the same time that it is exploring the new space, the spatial map remains stable for days and the animal readily remembers a task based on knowledge of that environment. (p. 312)
In The Principles of Psychology [1890] William James pointed out that there is more than one form of attention. There are at least two types: involuntary and voluntary. Involuntary attention is supported by automatic neural processes, and is particularly evident in implicit memory. Involuntary attention is activated by a property of the external world — of the stimulus — and it is captured, according to James, by “big things, bright things, moving things, or blood.” Voluntary attention, on the other hand, such as paying attention to the road and traffic while driving, is a specific feature of explicit memory and arises from the internal need to process stimuli that are not automatically salient. (p. 313)
One of the key differences between [involuntary and voluntary attention] is not the absence or presence of salience, but whether the signal of salience is perceived consciously. Studies also suggest that, as James had argued, the determining factor in whether memory is implicit or explicit is the manner in which the attentional signal for salience is recruited. (pp. 313-314)
In both types of memory, conversion of short-term to long-term memory requires the activation of genes, and in each case modulatory transmitters appear to carry an attentional signal marking the importance of a stimulus. In response to that signal, genes are turned on and proteins are produced and sent to all the synapses. But these signals of salience are called up in fundamentally different ways for the implicit memory and for the explicit memory required to form the spatial map in the mouse.
(p. 314)In the implicit memory storage, the attentional signal is recruited involuntarily (reflexively), from the bottom up: the sensory neurons of the tail, activated by a shock, act directly on the cells that release serotonin. In spatial memory, dopamine [a neurotransmitter] appears to be recruited voluntarily, from the top down: the cerebral cortex activates the cells that release dopamine, and dopamine modulates activity in the hippocampus. (p. 314)
The “presence of salience” is a higher-level form of what Gregory Bateson called “news of difference”. In Symbolic Modelling we are always modelling for “salience” (otherwise known as ‘sorting for significance’). And in particular “stimuli that are not automatically salient.” Kandel suggests attending to salient features is “a determining factor in the conversion of short-term to long-term memory.” From this we infer that inviting clients to voluntarily attend to, and maintain attention on salient features is a determining factor in encouraging changes that happen in the session to become long-term memories which are then involuntarily (i.e. unconsciously) “recruited” when needed in the future.
This suggests an important role for us as facilitators. We can pay attention to what the client finds salient and honour or “bless” that, and we can direct attention to salience that the client pays scant attention to. Some events and processes that do not automatically recruit voluntary attention are:
- Effects, and especially effects of effects, etc.
- “Weak links” (see our notes Thinking Networks)
- Relationship between nonadjacent items in terms of space, time, form and order (see our article on Proximity and Meaning)
- Scaling (see our article Big Fish in a Small Pond)
- Perceiver and perspective (see the article The Perceivers)
- Uncomfortable/unpleasant sensations
- “Inconvenient truths” (see our article, Self-Deception, Self-Delusion, self-Denial)
- Small changes
- Slow changes (especially those with a long “lag” between cause and effect, see Peter Senge, The Fifth Discipline)
3. The role of long-term potentiation
Since space involves information acquired through several sensory modalities, it raises the questions: How is the spatial map established? And, once established, how is it maintained?
The spatial map of even a simple locale does not form instantaneously but over ten to fifteen minutes of the rat’s entrance into the new environment. This suggests that the formation of the map is a learning process; practice makes perfect also for space. Under optimal circumstances this map remains stable for weeks or even months, much like a memory process. (p. 309)
What is true for rats seems true for many humans too. It often takes 10-15 minutes for a Clean Space network to begin to form.
The first clue to the answers emerged in 1973 when Terje Lomo and Tim Bliss discovered that the neuronal pathways leading to the hippocampus of rabbits can be strengthened by a brief burst of neuronal activity. Lomo and Bliss called this form of synaptic facilitation long-term potentiation. It soon emerged that long-term potentiation occurs in all three of the pathways within the hippocampus and describes a family of slightly different mechanisms, each of which increases the strength of the synapse in response to different rates and patterns of stimulation. Moreover there is a distinction between the processes involved in acquiring the map (and holding on to it for a few hours) and maintaining the map in stable form for the long term.
(pp. 282-283 and p. 310)Key molecules [are] involved in long-term potentiation. [For example,] glutamate, a common amino acid, acts on two different types of receptors in the hippocampus, the AMPA [alpha-amino-3-hydroxy-5-methylisoxazole-4- propionic acid] receptor and the NMDA [N-methyl-D-aspartatic acid] receptor. The AMPA receptor mediates normal synaptic transmission and responds to an individual action potential in the pre synaptic neuron. The NMDA receptor, on the other hand, responds only to extraordinarily rapid trains of stimuli and is required for long-term potentiation. NMDA receptors can transmit the electrical signal of the synaptic potential into a biochemical signal. These biochemical reactions are important because they trigger molecular signals that can be broadcast throughout the cell and thus contribute to long-lasting synaptic modifications. (pp. 283-284)The analysis of how the NMDA receptor functions showed that it acts as a coincidence detector. It allows calcium ions to flow through its channel if and only if it detects the coincidence of two neural events, one pre synaptic and the other post synaptic. In response to certain patterns of stimulation, the post synaptic cell also sends a signal back to the pre synaptic cell calling for more glutamate. [These] repeated trains of electrical stimulation produce a late phase of long-term potentiation that lasts for more than a day. (p. 284 and pp. 292-293)
Long-term potentiation involves a self-sustaining feedback loop: a post synaptic cell triggers a pre synaptic cell to produce more glutamate which triggers the post synaptic cell and so on. The effect is long-term memory.
In Symbolic Modelling the Maturing a Change process may work in a similar fashion. The repeated “potentiation” of new symbols, functions or relationships helps to consolidate the changed Landscape. The above suggests this process would be aided by facilitating the client to attend to the reorganised spatial configuration of symbols or spaces. Ways to do this are:
- Direct questions to any changes in spatial relations, e.g. distance, angle, boundary, etc.
- Ask questions of the changed Landscape which require spatial “cognitive sensibilities”,
e.g.
- What’s between X and Y? (and other specialised questions about space)
- When X is there, what happens to Y?
- What does X know from there about Y over there? (A Clean Space question which can also be used within a metaphor landscape.)
- Physicalise the landscape and visit different symbols noting their changed spatial relationships and perspectives.
- If a client using Clean Space moves some of their spaces, or discovers new spaces, at the end of the session the client can be invited to “walk to each space … and between the spaces … and round the periphery of the spaces … and notice where each space is in relation to the other spaces … and whether the spaces have a shape or a configuration.”
Other Thoughts
There are two common ways to do ‘second position’ NLP modelling, i.e. imagine being someone else. Either:
a. Move your ‘I’ into their body (e.g. Steve Gilligan’s ‘deep trance identification’ modelling)
or
b. Move their ‘I’ into your body (Richard Bandler’s ‘Teach me how you do you’ modelling).
(a) Sees the external world as relative to the other person’s body and presumably uses some form of allocentric coordinates.
(b) Maintains the external world relative to your body and therefore uses egocentric coordinates.
And there is another way to model someone else’s perspective — the Tompkins and Lawley’s way to model symbolically as a the clean facilitator:
c. In Integral Spirituality, Ken Wilber calls this persepctive the “Outside view of the Inside of another’s I” because you imagine what another person is experiencing from their (Inside) perspective while maintaining your own (Outside) perspective. We think this is their egocentric perspective within your allocentric perspective.
To use a metaphor:
(a) is like sitting in the passenger seat of someone else’s car while they are driving and mimicking how the car is being driven.
(b) is like driving your own car following instructions from someone else as they tell you how they would drive it.
(c) is like driving your own car while your Sat Nav is showing you what someone else is experiencing while they are driving their own car.
The Neurobiology of Space - Exercises
Three exploratory exercises formed the context for The Developing Group day, 4 August, 2007
Exercise 1 – Modelling Spaces
In pairs – 20 minutes each way
Purpose:
For the client to self-model their perception of several ‘physical’ spaces.
For the facilitator to develop their skills at modelling and maintaining a person’s attention on spatial information.
Facilitator says:
1. Where do you want to be and where do you want me to be? (Client locates both)
2. What spaces do you notice?
3. Select 3 of those spaces spaces.
4. Facilitate the person to describe the attributes of each of the 3 spaces.
5. And then, what’s the same and what’s different about the 3 spaces?
Things for the facilitator to notice during the exercise:
What kind of places are called ‘spaces’?
As we cannot directly perceive space, we have to compute it, what characteristics are used to categorise spaces?
When does a person’s description became less external-sensory and more imaginary-metaphorical, e.g.
b) It seems to have a direction.
c) I can imagine myself standing in that space.
Exercise 2 – Blind Clean Space
In pairs – 30 minutes each way.
Purpose:
Activity
- Represent a desired outcome (=B).
- Place B where it needs to be.
- Put on blindfold.
- Locate starting position (=A).
- Facilitator marks with a post-it note each space (about six) visited by the client and in which direction they were facing.
At the end the client removes blindfold and notices the configuration of spaces.
The words in red below are the changes from the original process described in our article Clean Space: Modelling human perception through emergence
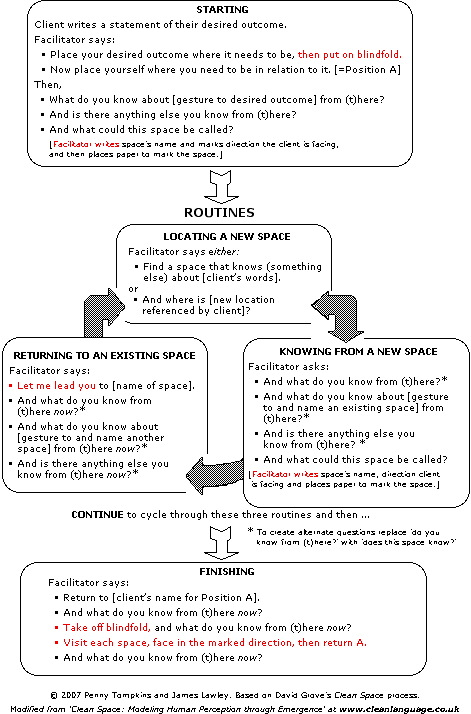
Exercise 3 – Spaces Between
In pairs – 30 minutes each way.
Acknowledgement: We designed this activity using a mixture of processes originated by David Grove – Clean Language, Clean Space, Clean Worlds and Emergent Knowledge. Articles on these subjects can be found in the menus on the left of this page.
Activity
On your own:
1. Identify a topic of interest or a desired outcome.
Represent it on a piece of paper (label it B).
2. What do you know about that B?
Represent that knowing on another piece of paper. (label it A).
3. Ask yourself six times: And what else do I know about B?
i.e. identify six other knowings and put each of those on separate pieces of paper
(number them 1 to 6).
You should now have 8 pieces of paper.
With a partner:
NB: This exercise is about the ‘space between’ and not about the content on the paper.
4. Place B where it needs to be.
Place yourself where you need to be in relation to B.
Put A where you are.
Place the six other knowings where they need to be in relation to A and B.
5. Starting at A, visit each space in turn and notice the extent of each space.
(Facilitator to note the client’s words that describe the space between,
e.g. ‘gap’, ‘shared boundary’, ‘edge’, ‘overlap’, etc.)
6. Return to A and select a space-between.
Identify the attributes of the space-between selected,
e.g.
- And does that … have a size or a shape? (… is a space between word)
- And what’s on either side of that …?
- And how far does the ‘overlap’ overlap?
- And how far does … extend?
- And what’s behind …?
- And what’s between … and …?
7. Select another ‘space between’ and repeat Step 6 several times
8. End by returning to A and asking:
- And what do you know now?
- And is there anything about these spaces that needs to be different?
If yes (and after they have made the alteration):
- And what do you know now?
Postscript
Follow-up articles:
How the Brain Feels: Emotion and Cognition in Neurolinguitsic Psychotherapy (2002-2007), Philip Harland’s excellent five-part paper.
Attending to Salience (2009). Penny Tompkins and my article goes into some depth on what guides our line of questioning and gives the session its sense of directional flow.
Pointing to a New Modelling Perspective (2012). My paper provides a different (and I now think more appropriate) metaphor for the clean facilitator’s perspective discussed above.
A Modeller’s Perspective (2014). My blog exploring six perspectives for modelling (the three mentioned above and three others).
Clean Space revisited (2009) With the benefit of seven years of experience and experimenting, an updated version of our first model.
27 Jan 2026
The article was modified to clarify the distinction between ego/allocentric coordinates and route/survey perspectives.
My supposition that, phenomenologically, egocentric coordinates are always taken into account, even when allocentric coordinates are used was verified by this research:
Filimon F (2015). Are All SpatialReference Frames Egocentric? Reinterpreting Evidence for Allocentric, Object-Centered, or World-Centered Reference Frames. Frontiers in Human Neuroscience 9:648. doi: 10.3389/fnhum.2015.0064.





